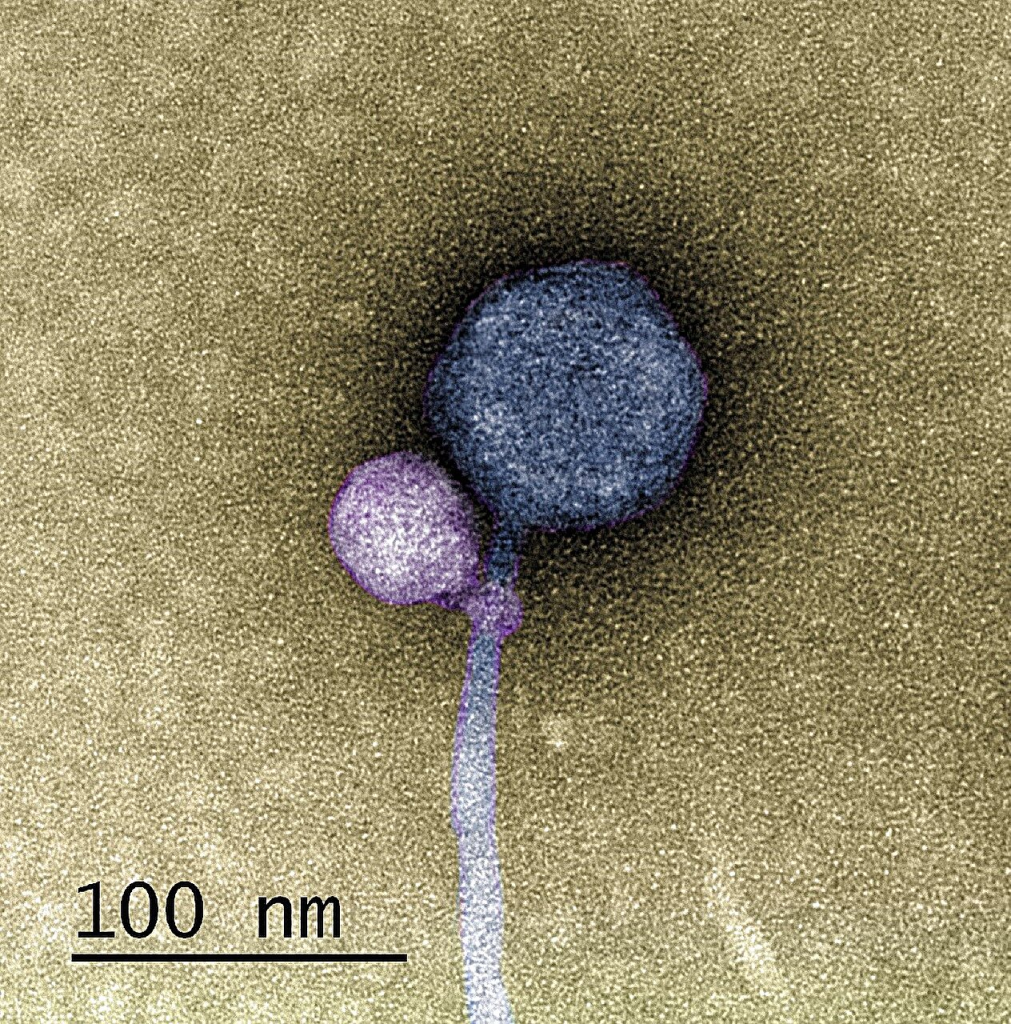
Discovery: A virus that attaches to another virus November 3, 2023 In a groundbreaking discovery that began as a typical research project, a team from the University of Maryland, Baltimore County (UMBC) uncovered a viral
Phage Facilitates Nitrogen Metabolism in Newly Discovered Deep-sea Bacteria September 5, 2023 Scientists have made important progress in understanding deep-sea Planctomycetes bacteria. These bacteria are found in various ecosystems and play a vital role in global
Inoviridae (Filamentous phages): Bacteriophages with a unique replication cycle July 3, 2022 Bacteriophage is a term used to describe viruses that infect bacteria and archaea. While most phages are 'micro predators' that kill their bacterial hosts,
What do Bacteriophage Diagrams Look Like? (Morphological classification of bacteriophages) May 4, 2022 What do bacteriophages look like? In both academic and non-academic contexts around the world, a well-shaped particle with a clearly separated head, tail (neck,